User login
Charles Darwin once said that apes “...are quite unable, as I have myself seen, to throw a stone with precision”.1 Yet humans can throw with precision and speed, a skill that likely had significant advantages: throwing can affect change at a distance—something few species can do. Throwing can provide protection against predators and can allow for predation for food resources. Throwing would be important in contesting other hominids for scarce resources. As such, throwing has been critically important in human evolution and likely is a skill that has been promoted through natural selection.2-5
In the orthopedic literature, most published work on throwing will ask proximate questions: “how, what, who, when, and where?” Evolutionary biologists are concerned with ultimate questions6,7: “why?” Asking ultimate questions provides insight into how a behavior might offer advantages under natural selection, which can then improve our understanding of the proximate questions for that behavior.
With regard to the shoulder, a number of mysteries exist that, to date, proximate studies have not been able to solve. This article argues that the human shoulder has evolved for throwing and by using this frame of reference, many of the mysteries surrounding the anatomy of the shoulder can be understood.
Pitching Kinematics
The mechanics of pitching have been analyzed extensively. Fleisig and colleagues8 performed kinematic and electromyographic analyses of pitchers to identify the critical moments of pitching (defined as where the forces are highest and injury is most likely going to occur). They found 2 moments where the forces about the shoulder are highest during pitching: the late cocking phase (defined by the point where the humerus reaches maximal external rotation); and the early deceleration phase (defined by the point when the ball is released). If throwing is important in natural selection of humans, then the shoulder anatomy should be optimized to withstand the forces generated in these positions.
Late Cocking Phase of Throwing
The early phases of throwing are attempting to maximize external rotation of the abducted arm as the velocity of the pitched ball correlates to the amount of external rotation achieved.9-11 In this position, kinetic energy in external rotation is stored and converted into kinetic energy in internal rotation.12 The position of the shoulder during late cocking is 94 ± 21° of thoracohumeral abduction, 11 ± 11° of horizontal adduction, and a remarkable 165 ± 11° of thoracohumeral external rotation (Figure 1).8
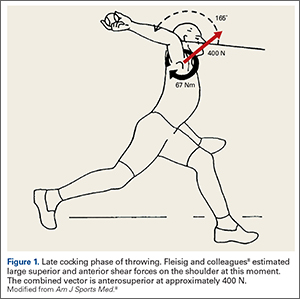
Fleisig and colleagues8 estimated the torque and forces about the shoulder, which are quite high for joint compression (480 ± 130 N). They also analyzed the shear forces and while trying to describe the origin of superior labrum anterior to posterior (SLAP) lesions and anterior labral tears, broke down the major shear vector into an anterior force vector (310 ± 100 N) and a superior force (250 ± 80 N).8 Note that the resulting shear vector is in an anterosuperior direction and is approximately 400 N.
Early Deceleration Phase of Throwing
Interestingly, the position of the humerus during this critical moment of throwing is not much different than the position during the late cocking phase of throwing, with 93 ± 10° of thoracohumeral abduction, 6 ± 8° of horizontal adduction.8 The major difference in the position of the arm is found in the amount of thoracohumeral rotation, which is now 64 ± 35° of external rotation (Figure 2).8
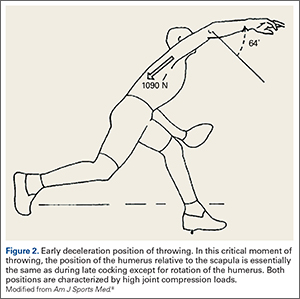
The forces in early deceleration are tremendous, with an estimated 1090 ± 110 N joint compression force, and an anteroinferior shear force of approximately 130 N.8
Clearly, if throwing is an important skill in human evolution, adaptations must exist in the shoulder to withstand the high forces in these 2 critical moments of throwing.
Solving Mysteries of Shoulder Anatomy in the Context of Throwing
There are many anatomic features of the shoulder that remain poorly understood. These include the alignment of the glenohumeral joint, the function of the glenohumeral ligaments, the function of the coracoacromial ligament, the depression of the human greater tuberosity, and the nature and function of the very tendinous subscapularis and long head of the biceps. These mysteries of the human shoulder can be solved if one considers the hypothesis that the shoulder has evolved to throw.
Glenohumeral Joint Alignment
The cartilage of the humeral head is thickest at its center, and thinnest at the periphery (Figure 3A).13,14 Conversely, the cartilage of the glenoid is thinnest at the fovea and thickest in the periphery (Figure 3B).14 It seems obvious that in order to maximally distribute high loads across this joint, the center of the humeral head should rest in the center of the glenoid. Interestingly, this does not occur during most positions of the shoulder. When upright, the center of the humeral head is directed above the glenoid in the coronal plane (Figure 3C). In order to align the glenohumeral joint optimally for the distribution of loads across the joint, the humerus must be abducted approximately 60° relative to the scapula. Assuming a 2:1 glenohumeral to scapulothoracic abduction for arm abduction relative to the thorax,15 this equates to approximately 90° of thoracohumeral abduction—the exact kinematic position of the shoulder during both critical moments of throwing (Figure 3D).
Function of the Glenohumeral Ligaments
The glenohumeral joint capsule has thickenings that help to stabilize the joint. The function of these glenohumeral ligaments has been evaluated biomechanically for their role in preventing translation and instability by a number of authors. The inferior glenohumeral ligament has classically been described as resisting anterior translation of the abducted arm.16 The coracohumeral ligament has been described as important to prevent inferior translation of the adducted arm.17
Interestingly, these ligaments are also the most important ligaments in resisting external rotation of the adducted arm.18 The dominant arm of throwing athletes has been shown to have increased inferior translation19 and increase external rotation.19-22 While the external rotation is partly related to bony adaptation,23,24 the ligamentous restraints to external rotation are likely under tremendous load, which may explain why Dr. Frank Jobe revolutionized the surgical treatment of the throwing athlete by performing an “instability” operation,25,26 as he believed these athletes had “subtle instability” that produced pain, but not symptoms of looseness.27
While these ligaments may exist in part to prevent translation and instability, current thinking suggests that “over-rotation” may lead to internal impingement and may be responsible for symptoms in the thrower’s shoulder,28 as SLAP lesions seem to occur easier with external rotation.29 Again, the importance of maximizing external rotation in throwing and the finding that this position is a critical moment with very high forces suggests that these ligaments may represent an adaptation to restrain external rotation while throwing.
Coracoacromial Ligament
The coracoacromial ligament is unique in that it connects 2 pieces of the same bone, and is only seen in hominids—not other primates.30 Its function has been debated for decades. This ligament is generally thought to limit superior translation of the humeral head,31,32 an effect that is critically important in patients with rotator cuff tears 33,34 Its importance is demonstrated by the fact that it seems to regenerate after it has been resected.35,36 Yet release or resection of this ligament has been a standard treatment for shoulder pain for decades.
Its function becomes clear if one examines the coracoacromial ligament with respect to the kinematics of throwing. As mentioned above, in the late cocking phase of throwing, tremendous shear forces exist in the shoulder. Fleisig and colleagues8 estimated a superior force of 250 ± 80 N, and an anterior shear force of 310 ± 100 N. While Fleisig and colleagues8 analyzed these shear forces with respect to the development of superior and anterior labral tears, it is important to note that these shear forces are vectors that should be combined. When one does this, it becomes apparent that in the late cocking phase of throwing there is shear force in an anterosuperior direction of approximately 400 N (Figure 1). The coracoacromial ligament is positioned to restrain this tremendous force. If throwing is an important adaptation in the evolution of humans, then the function of this ligament and its importance becomes clear.
Depressed Greater Tuberosity and the Pear-Shaped Glenoid
Compared to other primates, the greater tuberosity in humans sits significantly lower (Figure 4). This depression effectively decreases the moment arm of the muscle tendon unit, making the supraspinatus less powerful for raising the arm.37 In addition, by tenting the supraspinatus tendon over the humeral head, a watershed zone is created with decreased vascularity, which is thought to contribute to rotator cuff disease.38 What would be the advantage of the depressed tuberosity?
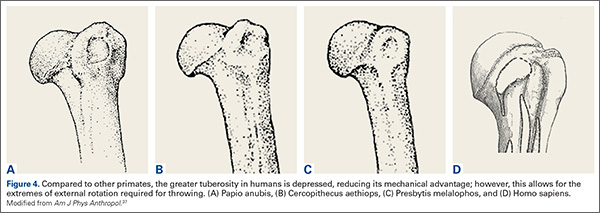
In primates, a lower tuberosity allows for more motion, particularly for arboreal travel.37 In order to throw with velocity, the humerus must achieve extremes of external rotation. A large tuberosity would limit external rotation of the abducted arm. Similarly, the pear-shaped glenoid cavity allows for the depressed tuberosity to achieve maximal external rotation. It is conceivable that a depressed greater tuberosity that allows for throwing would be an adaptation that could be favorable despite its proclivity toward rotator cuff disease in senescence.
Nature of the Subscapularis and the Role of the Long Head of the Biceps
The subscapularis is unique among rotator cuff muscles in that the upper two-thirds of the muscle is surprisingly tendinous.39 Why should this rotator cuff muscle have so much tendon material? Why is the tendon missing from the inferior one-third of the muscle? This situation is not optimal to prevent anterior glenohumeral instability, where inferior tendon material would be preferred.40
The function of the tendon of the long head of the biceps has long been debated and remains unclear.41-43 Cadaver experiments suggest the long head of the biceps provides glenohumeral joint stability in a variety of directions and positions, yet in vivo studies may not show this effect. Electromyography studies show little activity of the long head of the biceps with shoulder motion when the elbow is immobilized, leading some to suggest it is important as a passive restraint.43 This lack of understanding has led some to believe the biceps is not important and can be sacrificed without much concern.42,43
Again, these questions can be answered if one considers them in the context of throwing. At the point of maximal external rotation, the shoulder quickly moves from external rotation to internal rotation. This occurs by converting kinetic energy of external rotation into stored potential energy in the tissues. This energy is then converted into internal rotation. This elastic energy storage is critical for developing the necessary velocities to launch a projectile. While many structures are responsible for storing this energy,12 the subscapularis and long head of the biceps are particularly important. In fact, these 2 structures are important restraints to external rotation of the abducted arm–and become increasingly important with increased external rotation.45,46
One can think of the long head of the biceps as a spring (muscle), a cable (the long tendon), and a pulley (the bicipital groove). Similarly, one can consider the subscapularis as a similar structure, with the coracoid process serving as the pulley. In the late cocking phase of throwing, an interesting alignment occurs such that the pulleys (coracoid process and bicipital groove) are on opposite sides of the joint, providing glenohumeral joint stability. This system, with the inferior glenohumeral ligament (which is the primary restraint to external rotation of the abducted arm18), produces an incredibly stable envelope, preventing the humeral head from over-rotating and translating during the late cocking phase of throwing when the forces about the shoulder are extremely high. Because the muscles serve as springs, this system is also capable of storing kinetic energy during the late cocking phase of throwing and converting it into kinetic energy for internal rotation.
Summary
While throwing is not as critical to survival in today’s culture, the ability to throw was clearly an important adaptation in human evolution. With this in mind, we can approach human anatomy with this perspective, and in fact, many other lines of thinking suggest that throwing was important in the evolution of the hand,47 the brain,48 bipedalism,49 and even human society.50 The shoulder was highly influenced through natural selection to promote the throwing skill. With this perspective, many of the mysteries about the shoulder can be answered.
1. Darwin C. The Descent of Man, and Selection in Relation to Sex. 2nd ed. London, UK: John Murray; 1874:35.
2. Issac B. Throwing and human evolution. African Archeol Record. 1987;5:3-17.
3. Kirschann E. The human throw and a new model of hominid evolution (German). Homo. 1999;50(1):80-85.
4. Knusel CJ. The throwing hypothesis and hominid origins. Human Evolution. 1992;7(1):1-7.
5. Dunsworth H, Challis J, Walker A. The evolution of throwing: a new look at an old idea. Courier Forschungsinstitut Senckenberg. 2003;243:105-110.
6. Mayr E. Animal Species and Evolution. Cambridge, MA: Harvard University Press; 1963.
7. Tinbergen N. On the aims and methods of ethology. Zeitschrift für Tierpsychologie. 1963;20:410-433.
8. Fleisig GS, Andrews JR, Dillman CJ, Escamilla RF. Kinetics of baseball pitching with implications about injury mechanisms. Am J Sports Med. 1995;23(2):233-239.
9. Atwater AE. Biomechanics of overarm throwing movements and of throwing injuries. Exerc Sport Sci Rev. 1975;7:43-85.
10. Matsuo T, Escamila RF, Fleisig GS, Barrentine SW, Andrews JR. Comparison of kinematic and temporal parameters between different pitch velocity groups. J Appl Biomech. 2001;17:1-13.
11. Wang YT, Ford HT III, Ford HT Jr, Shin DM. Three-dimensional kinematic analysis of baseball pitching in acceleration phase. Percept Mot Skills. 1995;80:43-48.
12. Roach NT, Venkadesan M, Rainbow MJ, Lieberman DE. Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. Nature. 2013;498(7455):483-486.
13. Fox JA, Cole BJ, Romeo AA, et al. Articular cartilage thickness of the humeral head: an anatomic study. Orthopedics. 2008;31(3):216.
14. Zumstein V, Kraljevic M, Conzen A, Hoechel S, Müller-Gerbl M. Thickness distribution of the glenohumeral joint cartilage: a quantitative study using computed tomography. Surg Radiol Anat. 2014;36(4):327-331.
15. Inman VT, Saunders M, Abbott LC. Observations on the function of the shoulder joint. J Bone Joint Surg Am. 1944;26:1-30.
16. O’Brien SJ, Schwartz RS, Warren RF, Torzilli PA. Capsular restraints to anterior posterior motion of the abducted shoulder: A biomechanical study. J Shoulder Elbow Surg. 1995;4(4):298-308.
17. Warner JJ, Deng XH, Warren RF, Torzilli PA. Static capsuloligamentous restraints to superior-inferior translation of the glenohumeral joint. Am J Sports Med. 1992;20(6):675-685.
18. Kuhn JE, Bey MJ, Huston LJ, Blasier RB, Soslowsky LJ. Ligamentous restraints to external rotation of the humerus in the late-cocking phase of throwing. A cadaveric biomechanical investigation. Am J Sports Med. 2000;28(2):200-205.
19. Bigliani LU, Codd TP, Connor PM, Levine WN, Littlefield MA, Hershon SJ. Shoulder motion and laxity in the professional baseball player. Am J Sports Med. 1997;25(5):609-613.
20. Borsa PA, Dover GC, Wilk KE, Reinold MM. Glenohumeral range of motion and stiffness in professional baseball pitchers. Med Sci Sports Exerc. 2006;38(1):21-26.
21. Hurd WJ, Kaplan KM, Eiattrache NS, Jobe FW, Morrey BF, Kaufman KR. A profile of glenohumeral internal and external rotation motion in the uninjured high school baseball pitcher, part I: motion. J Athl Train. 2011;46(3):282-288.
22. Wilk KE, Macrina LC, Arrigo C. Passive range of motion characteristics in the overhead baseball pitcher and their implications for rehabilitation. Clin Orthop Relat Res. 2012;470(6):1586-1594.
23. Osbahr DC, Cannon DL, Speer KP. Retroversion of the humerus in the throwing shoulder of college baseball pitchers. Am J Sports Med. 2002;30(3):347-353.
24. Greenberg EM, Fernandez-Fernandez A, Lawrence JT, McClure P. The development of humeral retrotorsion and its relationship to throwing sports. Sports Health. 2015;7(6):489-496.
25. Jobe FW, Pink M. The athlete’s shoulder. J Hand Ther. 1994;7(2):107-110.
26. Montgomery WH 3rd, Jobe FW. Functional outcomes in athletes after modified anterior capsulolabral reconstruction. Am J Sports Med. 1994;22(3):352-358.
27. Jobe FW, Kvitne RS, Giangarra CE. Shoulder pain in the overhand or throwing athlete. The relationship of anterior instability and rotator cuff impingement. Orthop Rev. 1989;18(9):963-975.
28. Reinhold MM, Wilk KE, Dugas JR, Andrews JR. Chapter 11. Internal Impingement. In: Wilk K, Reinold MM, Andrews JR, eds. The Athlete’s Shoulder. 2nd ed. Philadelphia, PA: Churchill Livingstone Elsevier; 2009:126.
29. Kuhn JE, Lindholm SR, Huston LJ, Soslowsky LJ, Blasier RB. Failure of the biceps superior labral complex: a cadaveric biomechanical investigation comparing the late cocking and early deceleration positions of throwing. Arthroscopy. 2003;19(4):373-379.
30. Ciochon RL, Corruccini RS. The coraco-acromial ligament and projection index in man and other anthropoid primates. J Anat. 1977;124(Pt 3):627-632.
31. Moorman CT, Warren RF, Deng XH, Wickiewicz TL, Torzilli PA. Role of coracoacromial ligament and related structures in glenohumeral stability: a cadaveric study. J Surg Orthop Adv. 2012;21(4):210-217.
32. Su WR, Budoff JE, Luo ZP. The effect of coracoacromial ligament excision and acromioplasty on superior and anterosuperior glenohumeral stability. Arthroscopy. 2009;25(1):13-18.
33. Wellmann M, Petersen W, Zantop T, Schanz S, Raschke MJ, Hurschler C. Effect of coracoacromial ligament resection on glenohumeral stability under active muscle loading in an in vitro model. Arthroscopy. 2008;24(11):1258-1264.
34. Fagelman M, Sartori M, Freedman KB, Patwardhan AG, Carandang G, Marra G. Biomechanics of coracoacromial arch modification. J Shoulder Elbow Surg. 2007;16(1):101-116.
35. Bak K, Spring IB, Henderson IP. Re-formation of the coracoacromial ligament after open resection or arthroscopic release. J Shoulder Elbow Surg. 2000;9:289-293.
36. Levy O, Copeland SA. Regeneration of the coracoacromial ligament after acromioplasty and arthroscopic subacromial decompression. J Shoulder Elbow Surg. 2001;10(4):317-320.
37. Larson SG, Stern JT Jr. Role of supraspinatus in the quadrupedal locomotion of vervets (Cercopithecus aethiops): Implications for interpretation of humeral morphology. Am J Phys Anthropol. 1989;79(3):369-377.
38. Chansky HA, Iannotti JP. The vascularity of the rotator cuff. Clin Sports Med. 1991;10(4):807-822.
39. Klapper RC, Jobe FW, Matsuura P. Subscapularis muscle and its glenohumeral ligament-like bands. A histomorphologic study. Am J Sports Med. 1992;20(3):307-310.
40. Halder A, Zobitz ME, Schultz E, An KN. Structural properties of the subscapularis tendon. J Orthop Res. 2000;18(5):
829-834.
41. Elser F, Braun S, Dewing CB, Giphart JE, Millett PJ. Anatomy, function, injuries, and treatment of the long head of the biceps brachii tendon. Arthroscopy. 2011;27(4):581-592.
42. Pill SG, Walch G, Hawkins RJ, Kissenberth MJ. The role of the biceps tendon in massive rotator cuff tears. Instr Course Lect. 2012;61:113-120.
43. Krupp RJ, Kevern MA, Gaines MD, Kotara S, Singleton SB. Long head of the biceps tendon pain: differential diagnosis and treatment. J Orthop Sports Phys Ther. 2009;39(2):55-70.
44. Levy AS, Kelly BT, Lintner SA, Osbahr DC, Speer KP. Function of the long head of the biceps at the shoulder: electromyographic analysis. J Shoulder Elbow Surg. 2001;10(3):250-255.
45. Kuhn JE, Huston LJ, Soslowsky LJ, Shyr Y, Blasier RB. External rotation of the glenohumeral joint: ligament restraints and muscle effects in the neutral and abducted positions.
J Shoulder Elbow Surg. 2005;14(1 Suppl S):39S-48S.
46. McGarry MH, Nguyen ML, Quigley RJ, Hanypsiak B, Gupta R, Lee TQ. The effect of long and short head biceps loading on glenohumeral joint rotational range of motion and humeral head postion. Knee Surge Sports Traumatol Arthrosc. 2014 Sep 26. [Epub ahead of print].
47. Young RW. Evolution of the human hand: The role of throwing and clubbing. J Anat. 2003;202:165-174.
48. Calvin WH. Did throwing stones shape hominid brain evolution? Ethology and Sociobiology. 1982;3:115-124.
49. Fifer FC. The adoption of bipedalism by the hominids: A new hypothesis. Human Evolution. 1987;2(2):135-147.
50. Darlington PJ. Group selection, altruism, reinforcement, and throwing in human evolution. Proc Nat Acad Sci. 1973;72(9):3748-3752.
Charles Darwin once said that apes “...are quite unable, as I have myself seen, to throw a stone with precision”.1 Yet humans can throw with precision and speed, a skill that likely had significant advantages: throwing can affect change at a distance—something few species can do. Throwing can provide protection against predators and can allow for predation for food resources. Throwing would be important in contesting other hominids for scarce resources. As such, throwing has been critically important in human evolution and likely is a skill that has been promoted through natural selection.2-5
In the orthopedic literature, most published work on throwing will ask proximate questions: “how, what, who, when, and where?” Evolutionary biologists are concerned with ultimate questions6,7: “why?” Asking ultimate questions provides insight into how a behavior might offer advantages under natural selection, which can then improve our understanding of the proximate questions for that behavior.
With regard to the shoulder, a number of mysteries exist that, to date, proximate studies have not been able to solve. This article argues that the human shoulder has evolved for throwing and by using this frame of reference, many of the mysteries surrounding the anatomy of the shoulder can be understood.
Pitching Kinematics
The mechanics of pitching have been analyzed extensively. Fleisig and colleagues8 performed kinematic and electromyographic analyses of pitchers to identify the critical moments of pitching (defined as where the forces are highest and injury is most likely going to occur). They found 2 moments where the forces about the shoulder are highest during pitching: the late cocking phase (defined by the point where the humerus reaches maximal external rotation); and the early deceleration phase (defined by the point when the ball is released). If throwing is important in natural selection of humans, then the shoulder anatomy should be optimized to withstand the forces generated in these positions.
Late Cocking Phase of Throwing
The early phases of throwing are attempting to maximize external rotation of the abducted arm as the velocity of the pitched ball correlates to the amount of external rotation achieved.9-11 In this position, kinetic energy in external rotation is stored and converted into kinetic energy in internal rotation.12 The position of the shoulder during late cocking is 94 ± 21° of thoracohumeral abduction, 11 ± 11° of horizontal adduction, and a remarkable 165 ± 11° of thoracohumeral external rotation (Figure 1).8
Fleisig and colleagues8 estimated the torque and forces about the shoulder, which are quite high for joint compression (480 ± 130 N). They also analyzed the shear forces and while trying to describe the origin of superior labrum anterior to posterior (SLAP) lesions and anterior labral tears, broke down the major shear vector into an anterior force vector (310 ± 100 N) and a superior force (250 ± 80 N).8 Note that the resulting shear vector is in an anterosuperior direction and is approximately 400 N.
Early Deceleration Phase of Throwing
Interestingly, the position of the humerus during this critical moment of throwing is not much different than the position during the late cocking phase of throwing, with 93 ± 10° of thoracohumeral abduction, 6 ± 8° of horizontal adduction.8 The major difference in the position of the arm is found in the amount of thoracohumeral rotation, which is now 64 ± 35° of external rotation (Figure 2).8
The forces in early deceleration are tremendous, with an estimated 1090 ± 110 N joint compression force, and an anteroinferior shear force of approximately 130 N.8
Clearly, if throwing is an important skill in human evolution, adaptations must exist in the shoulder to withstand the high forces in these 2 critical moments of throwing.
Solving Mysteries of Shoulder Anatomy in the Context of Throwing
There are many anatomic features of the shoulder that remain poorly understood. These include the alignment of the glenohumeral joint, the function of the glenohumeral ligaments, the function of the coracoacromial ligament, the depression of the human greater tuberosity, and the nature and function of the very tendinous subscapularis and long head of the biceps. These mysteries of the human shoulder can be solved if one considers the hypothesis that the shoulder has evolved to throw.
Glenohumeral Joint Alignment
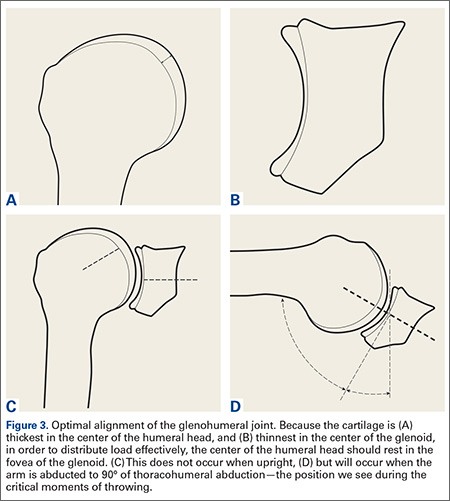
The cartilage of the humeral head is thickest at its center, and thinnest at the periphery (Figure 3A).13,14 Conversely, the cartilage of the glenoid is thinnest at the fovea and thickest in the periphery (Figure 3B).14 It seems obvious that in order to maximally distribute high loads across this joint, the center of the humeral head should rest in the center of the glenoid. Interestingly, this does not occur during most positions of the shoulder. When upright, the center of the humeral head is directed above the glenoid in the coronal plane (Figure 3C). In order to align the glenohumeral joint optimally for the distribution of loads across the joint, the humerus must be abducted approximately 60° relative to the scapula. Assuming a 2:1 glenohumeral to scapulothoracic abduction for arm abduction relative to the thorax,15 this equates to approximately 90° of thoracohumeral abduction—the exact kinematic position of the shoulder during both critical moments of throwing (Figure 3D).
Function of the Glenohumeral Ligaments
The glenohumeral joint capsule has thickenings that help to stabilize the joint. The function of these glenohumeral ligaments has been evaluated biomechanically for their role in preventing translation and instability by a number of authors. The inferior glenohumeral ligament has classically been described as resisting anterior translation of the abducted arm.16 The coracohumeral ligament has been described as important to prevent inferior translation of the adducted arm.17
Interestingly, these ligaments are also the most important ligaments in resisting external rotation of the adducted arm.18 The dominant arm of throwing athletes has been shown to have increased inferior translation19 and increase external rotation.19-22 While the external rotation is partly related to bony adaptation,23,24 the ligamentous restraints to external rotation are likely under tremendous load, which may explain why Dr. Frank Jobe revolutionized the surgical treatment of the throwing athlete by performing an “instability” operation,25,26 as he believed these athletes had “subtle instability” that produced pain, but not symptoms of looseness.27
While these ligaments may exist in part to prevent translation and instability, current thinking suggests that “over-rotation” may lead to internal impingement and may be responsible for symptoms in the thrower’s shoulder,28 as SLAP lesions seem to occur easier with external rotation.29 Again, the importance of maximizing external rotation in throwing and the finding that this position is a critical moment with very high forces suggests that these ligaments may represent an adaptation to restrain external rotation while throwing.
Coracoacromial Ligament
The coracoacromial ligament is unique in that it connects 2 pieces of the same bone, and is only seen in hominids—not other primates.30 Its function has been debated for decades. This ligament is generally thought to limit superior translation of the humeral head,31,32 an effect that is critically important in patients with rotator cuff tears 33,34 Its importance is demonstrated by the fact that it seems to regenerate after it has been resected.35,36 Yet release or resection of this ligament has been a standard treatment for shoulder pain for decades.
Its function becomes clear if one examines the coracoacromial ligament with respect to the kinematics of throwing. As mentioned above, in the late cocking phase of throwing, tremendous shear forces exist in the shoulder. Fleisig and colleagues8 estimated a superior force of 250 ± 80 N, and an anterior shear force of 310 ± 100 N. While Fleisig and colleagues8 analyzed these shear forces with respect to the development of superior and anterior labral tears, it is important to note that these shear forces are vectors that should be combined. When one does this, it becomes apparent that in the late cocking phase of throwing there is shear force in an anterosuperior direction of approximately 400 N (Figure 1). The coracoacromial ligament is positioned to restrain this tremendous force. If throwing is an important adaptation in the evolution of humans, then the function of this ligament and its importance becomes clear.
Depressed Greater Tuberosity and the Pear-Shaped Glenoid
Compared to other primates, the greater tuberosity in humans sits significantly lower (Figure 4). This depression effectively decreases the moment arm of the muscle tendon unit, making the supraspinatus less powerful for raising the arm.37 In addition, by tenting the supraspinatus tendon over the humeral head, a watershed zone is created with decreased vascularity, which is thought to contribute to rotator cuff disease.38 What would be the advantage of the depressed tuberosity?
In primates, a lower tuberosity allows for more motion, particularly for arboreal travel.37 In order to throw with velocity, the humerus must achieve extremes of external rotation. A large tuberosity would limit external rotation of the abducted arm. Similarly, the pear-shaped glenoid cavity allows for the depressed tuberosity to achieve maximal external rotation. It is conceivable that a depressed greater tuberosity that allows for throwing would be an adaptation that could be favorable despite its proclivity toward rotator cuff disease in senescence.
Nature of the Subscapularis and the Role of the Long Head of the Biceps
The subscapularis is unique among rotator cuff muscles in that the upper two-thirds of the muscle is surprisingly tendinous.39 Why should this rotator cuff muscle have so much tendon material? Why is the tendon missing from the inferior one-third of the muscle? This situation is not optimal to prevent anterior glenohumeral instability, where inferior tendon material would be preferred.40
The function of the tendon of the long head of the biceps has long been debated and remains unclear.41-43 Cadaver experiments suggest the long head of the biceps provides glenohumeral joint stability in a variety of directions and positions, yet in vivo studies may not show this effect. Electromyography studies show little activity of the long head of the biceps with shoulder motion when the elbow is immobilized, leading some to suggest it is important as a passive restraint.43 This lack of understanding has led some to believe the biceps is not important and can be sacrificed without much concern.42,43
Again, these questions can be answered if one considers them in the context of throwing. At the point of maximal external rotation, the shoulder quickly moves from external rotation to internal rotation. This occurs by converting kinetic energy of external rotation into stored potential energy in the tissues. This energy is then converted into internal rotation. This elastic energy storage is critical for developing the necessary velocities to launch a projectile. While many structures are responsible for storing this energy,12 the subscapularis and long head of the biceps are particularly important. In fact, these 2 structures are important restraints to external rotation of the abducted arm–and become increasingly important with increased external rotation.45,46
One can think of the long head of the biceps as a spring (muscle), a cable (the long tendon), and a pulley (the bicipital groove). Similarly, one can consider the subscapularis as a similar structure, with the coracoid process serving as the pulley. In the late cocking phase of throwing, an interesting alignment occurs such that the pulleys (coracoid process and bicipital groove) are on opposite sides of the joint, providing glenohumeral joint stability. This system, with the inferior glenohumeral ligament (which is the primary restraint to external rotation of the abducted arm18), produces an incredibly stable envelope, preventing the humeral head from over-rotating and translating during the late cocking phase of throwing when the forces about the shoulder are extremely high. Because the muscles serve as springs, this system is also capable of storing kinetic energy during the late cocking phase of throwing and converting it into kinetic energy for internal rotation.
Summary
While throwing is not as critical to survival in today’s culture, the ability to throw was clearly an important adaptation in human evolution. With this in mind, we can approach human anatomy with this perspective, and in fact, many other lines of thinking suggest that throwing was important in the evolution of the hand,47 the brain,48 bipedalism,49 and even human society.50 The shoulder was highly influenced through natural selection to promote the throwing skill. With this perspective, many of the mysteries about the shoulder can be answered.
Charles Darwin once said that apes “...are quite unable, as I have myself seen, to throw a stone with precision”.1 Yet humans can throw with precision and speed, a skill that likely had significant advantages: throwing can affect change at a distance—something few species can do. Throwing can provide protection against predators and can allow for predation for food resources. Throwing would be important in contesting other hominids for scarce resources. As such, throwing has been critically important in human evolution and likely is a skill that has been promoted through natural selection.2-5
In the orthopedic literature, most published work on throwing will ask proximate questions: “how, what, who, when, and where?” Evolutionary biologists are concerned with ultimate questions6,7: “why?” Asking ultimate questions provides insight into how a behavior might offer advantages under natural selection, which can then improve our understanding of the proximate questions for that behavior.
With regard to the shoulder, a number of mysteries exist that, to date, proximate studies have not been able to solve. This article argues that the human shoulder has evolved for throwing and by using this frame of reference, many of the mysteries surrounding the anatomy of the shoulder can be understood.
Pitching Kinematics
The mechanics of pitching have been analyzed extensively. Fleisig and colleagues8 performed kinematic and electromyographic analyses of pitchers to identify the critical moments of pitching (defined as where the forces are highest and injury is most likely going to occur). They found 2 moments where the forces about the shoulder are highest during pitching: the late cocking phase (defined by the point where the humerus reaches maximal external rotation); and the early deceleration phase (defined by the point when the ball is released). If throwing is important in natural selection of humans, then the shoulder anatomy should be optimized to withstand the forces generated in these positions.
Late Cocking Phase of Throwing
The early phases of throwing are attempting to maximize external rotation of the abducted arm as the velocity of the pitched ball correlates to the amount of external rotation achieved.9-11 In this position, kinetic energy in external rotation is stored and converted into kinetic energy in internal rotation.12 The position of the shoulder during late cocking is 94 ± 21° of thoracohumeral abduction, 11 ± 11° of horizontal adduction, and a remarkable 165 ± 11° of thoracohumeral external rotation (Figure 1).8
Fleisig and colleagues8 estimated the torque and forces about the shoulder, which are quite high for joint compression (480 ± 130 N). They also analyzed the shear forces and while trying to describe the origin of superior labrum anterior to posterior (SLAP) lesions and anterior labral tears, broke down the major shear vector into an anterior force vector (310 ± 100 N) and a superior force (250 ± 80 N).8 Note that the resulting shear vector is in an anterosuperior direction and is approximately 400 N.
Early Deceleration Phase of Throwing
Interestingly, the position of the humerus during this critical moment of throwing is not much different than the position during the late cocking phase of throwing, with 93 ± 10° of thoracohumeral abduction, 6 ± 8° of horizontal adduction.8 The major difference in the position of the arm is found in the amount of thoracohumeral rotation, which is now 64 ± 35° of external rotation (Figure 2).8
The forces in early deceleration are tremendous, with an estimated 1090 ± 110 N joint compression force, and an anteroinferior shear force of approximately 130 N.8
Clearly, if throwing is an important skill in human evolution, adaptations must exist in the shoulder to withstand the high forces in these 2 critical moments of throwing.
Solving Mysteries of Shoulder Anatomy in the Context of Throwing
There are many anatomic features of the shoulder that remain poorly understood. These include the alignment of the glenohumeral joint, the function of the glenohumeral ligaments, the function of the coracoacromial ligament, the depression of the human greater tuberosity, and the nature and function of the very tendinous subscapularis and long head of the biceps. These mysteries of the human shoulder can be solved if one considers the hypothesis that the shoulder has evolved to throw.
Glenohumeral Joint Alignment
The cartilage of the humeral head is thickest at its center, and thinnest at the periphery (Figure 3A).13,14 Conversely, the cartilage of the glenoid is thinnest at the fovea and thickest in the periphery (Figure 3B).14 It seems obvious that in order to maximally distribute high loads across this joint, the center of the humeral head should rest in the center of the glenoid. Interestingly, this does not occur during most positions of the shoulder. When upright, the center of the humeral head is directed above the glenoid in the coronal plane (Figure 3C). In order to align the glenohumeral joint optimally for the distribution of loads across the joint, the humerus must be abducted approximately 60° relative to the scapula. Assuming a 2:1 glenohumeral to scapulothoracic abduction for arm abduction relative to the thorax,15 this equates to approximately 90° of thoracohumeral abduction—the exact kinematic position of the shoulder during both critical moments of throwing (Figure 3D).
Function of the Glenohumeral Ligaments
The glenohumeral joint capsule has thickenings that help to stabilize the joint. The function of these glenohumeral ligaments has been evaluated biomechanically for their role in preventing translation and instability by a number of authors. The inferior glenohumeral ligament has classically been described as resisting anterior translation of the abducted arm.16 The coracohumeral ligament has been described as important to prevent inferior translation of the adducted arm.17
Interestingly, these ligaments are also the most important ligaments in resisting external rotation of the adducted arm.18 The dominant arm of throwing athletes has been shown to have increased inferior translation19 and increase external rotation.19-22 While the external rotation is partly related to bony adaptation,23,24 the ligamentous restraints to external rotation are likely under tremendous load, which may explain why Dr. Frank Jobe revolutionized the surgical treatment of the throwing athlete by performing an “instability” operation,25,26 as he believed these athletes had “subtle instability” that produced pain, but not symptoms of looseness.27
While these ligaments may exist in part to prevent translation and instability, current thinking suggests that “over-rotation” may lead to internal impingement and may be responsible for symptoms in the thrower’s shoulder,28 as SLAP lesions seem to occur easier with external rotation.29 Again, the importance of maximizing external rotation in throwing and the finding that this position is a critical moment with very high forces suggests that these ligaments may represent an adaptation to restrain external rotation while throwing.
Coracoacromial Ligament
The coracoacromial ligament is unique in that it connects 2 pieces of the same bone, and is only seen in hominids—not other primates.30 Its function has been debated for decades. This ligament is generally thought to limit superior translation of the humeral head,31,32 an effect that is critically important in patients with rotator cuff tears 33,34 Its importance is demonstrated by the fact that it seems to regenerate after it has been resected.35,36 Yet release or resection of this ligament has been a standard treatment for shoulder pain for decades.
Its function becomes clear if one examines the coracoacromial ligament with respect to the kinematics of throwing. As mentioned above, in the late cocking phase of throwing, tremendous shear forces exist in the shoulder. Fleisig and colleagues8 estimated a superior force of 250 ± 80 N, and an anterior shear force of 310 ± 100 N. While Fleisig and colleagues8 analyzed these shear forces with respect to the development of superior and anterior labral tears, it is important to note that these shear forces are vectors that should be combined. When one does this, it becomes apparent that in the late cocking phase of throwing there is shear force in an anterosuperior direction of approximately 400 N (Figure 1). The coracoacromial ligament is positioned to restrain this tremendous force. If throwing is an important adaptation in the evolution of humans, then the function of this ligament and its importance becomes clear.
Depressed Greater Tuberosity and the Pear-Shaped Glenoid
Compared to other primates, the greater tuberosity in humans sits significantly lower (Figure 4). This depression effectively decreases the moment arm of the muscle tendon unit, making the supraspinatus less powerful for raising the arm.37 In addition, by tenting the supraspinatus tendon over the humeral head, a watershed zone is created with decreased vascularity, which is thought to contribute to rotator cuff disease.38 What would be the advantage of the depressed tuberosity?
In primates, a lower tuberosity allows for more motion, particularly for arboreal travel.37 In order to throw with velocity, the humerus must achieve extremes of external rotation. A large tuberosity would limit external rotation of the abducted arm. Similarly, the pear-shaped glenoid cavity allows for the depressed tuberosity to achieve maximal external rotation. It is conceivable that a depressed greater tuberosity that allows for throwing would be an adaptation that could be favorable despite its proclivity toward rotator cuff disease in senescence.
Nature of the Subscapularis and the Role of the Long Head of the Biceps
The subscapularis is unique among rotator cuff muscles in that the upper two-thirds of the muscle is surprisingly tendinous.39 Why should this rotator cuff muscle have so much tendon material? Why is the tendon missing from the inferior one-third of the muscle? This situation is not optimal to prevent anterior glenohumeral instability, where inferior tendon material would be preferred.40
The function of the tendon of the long head of the biceps has long been debated and remains unclear.41-43 Cadaver experiments suggest the long head of the biceps provides glenohumeral joint stability in a variety of directions and positions, yet in vivo studies may not show this effect. Electromyography studies show little activity of the long head of the biceps with shoulder motion when the elbow is immobilized, leading some to suggest it is important as a passive restraint.43 This lack of understanding has led some to believe the biceps is not important and can be sacrificed without much concern.42,43
Again, these questions can be answered if one considers them in the context of throwing. At the point of maximal external rotation, the shoulder quickly moves from external rotation to internal rotation. This occurs by converting kinetic energy of external rotation into stored potential energy in the tissues. This energy is then converted into internal rotation. This elastic energy storage is critical for developing the necessary velocities to launch a projectile. While many structures are responsible for storing this energy,12 the subscapularis and long head of the biceps are particularly important. In fact, these 2 structures are important restraints to external rotation of the abducted arm–and become increasingly important with increased external rotation.45,46
One can think of the long head of the biceps as a spring (muscle), a cable (the long tendon), and a pulley (the bicipital groove). Similarly, one can consider the subscapularis as a similar structure, with the coracoid process serving as the pulley. In the late cocking phase of throwing, an interesting alignment occurs such that the pulleys (coracoid process and bicipital groove) are on opposite sides of the joint, providing glenohumeral joint stability. This system, with the inferior glenohumeral ligament (which is the primary restraint to external rotation of the abducted arm18), produces an incredibly stable envelope, preventing the humeral head from over-rotating and translating during the late cocking phase of throwing when the forces about the shoulder are extremely high. Because the muscles serve as springs, this system is also capable of storing kinetic energy during the late cocking phase of throwing and converting it into kinetic energy for internal rotation.
Summary
While throwing is not as critical to survival in today’s culture, the ability to throw was clearly an important adaptation in human evolution. With this in mind, we can approach human anatomy with this perspective, and in fact, many other lines of thinking suggest that throwing was important in the evolution of the hand,47 the brain,48 bipedalism,49 and even human society.50 The shoulder was highly influenced through natural selection to promote the throwing skill. With this perspective, many of the mysteries about the shoulder can be answered.
1. Darwin C. The Descent of Man, and Selection in Relation to Sex. 2nd ed. London, UK: John Murray; 1874:35.
2. Issac B. Throwing and human evolution. African Archeol Record. 1987;5:3-17.
3. Kirschann E. The human throw and a new model of hominid evolution (German). Homo. 1999;50(1):80-85.
4. Knusel CJ. The throwing hypothesis and hominid origins. Human Evolution. 1992;7(1):1-7.
5. Dunsworth H, Challis J, Walker A. The evolution of throwing: a new look at an old idea. Courier Forschungsinstitut Senckenberg. 2003;243:105-110.
6. Mayr E. Animal Species and Evolution. Cambridge, MA: Harvard University Press; 1963.
7. Tinbergen N. On the aims and methods of ethology. Zeitschrift für Tierpsychologie. 1963;20:410-433.
8. Fleisig GS, Andrews JR, Dillman CJ, Escamilla RF. Kinetics of baseball pitching with implications about injury mechanisms. Am J Sports Med. 1995;23(2):233-239.
9. Atwater AE. Biomechanics of overarm throwing movements and of throwing injuries. Exerc Sport Sci Rev. 1975;7:43-85.
10. Matsuo T, Escamila RF, Fleisig GS, Barrentine SW, Andrews JR. Comparison of kinematic and temporal parameters between different pitch velocity groups. J Appl Biomech. 2001;17:1-13.
11. Wang YT, Ford HT III, Ford HT Jr, Shin DM. Three-dimensional kinematic analysis of baseball pitching in acceleration phase. Percept Mot Skills. 1995;80:43-48.
12. Roach NT, Venkadesan M, Rainbow MJ, Lieberman DE. Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. Nature. 2013;498(7455):483-486.
13. Fox JA, Cole BJ, Romeo AA, et al. Articular cartilage thickness of the humeral head: an anatomic study. Orthopedics. 2008;31(3):216.
14. Zumstein V, Kraljevic M, Conzen A, Hoechel S, Müller-Gerbl M. Thickness distribution of the glenohumeral joint cartilage: a quantitative study using computed tomography. Surg Radiol Anat. 2014;36(4):327-331.
15. Inman VT, Saunders M, Abbott LC. Observations on the function of the shoulder joint. J Bone Joint Surg Am. 1944;26:1-30.
16. O’Brien SJ, Schwartz RS, Warren RF, Torzilli PA. Capsular restraints to anterior posterior motion of the abducted shoulder: A biomechanical study. J Shoulder Elbow Surg. 1995;4(4):298-308.
17. Warner JJ, Deng XH, Warren RF, Torzilli PA. Static capsuloligamentous restraints to superior-inferior translation of the glenohumeral joint. Am J Sports Med. 1992;20(6):675-685.
18. Kuhn JE, Bey MJ, Huston LJ, Blasier RB, Soslowsky LJ. Ligamentous restraints to external rotation of the humerus in the late-cocking phase of throwing. A cadaveric biomechanical investigation. Am J Sports Med. 2000;28(2):200-205.
19. Bigliani LU, Codd TP, Connor PM, Levine WN, Littlefield MA, Hershon SJ. Shoulder motion and laxity in the professional baseball player. Am J Sports Med. 1997;25(5):609-613.
20. Borsa PA, Dover GC, Wilk KE, Reinold MM. Glenohumeral range of motion and stiffness in professional baseball pitchers. Med Sci Sports Exerc. 2006;38(1):21-26.
21. Hurd WJ, Kaplan KM, Eiattrache NS, Jobe FW, Morrey BF, Kaufman KR. A profile of glenohumeral internal and external rotation motion in the uninjured high school baseball pitcher, part I: motion. J Athl Train. 2011;46(3):282-288.
22. Wilk KE, Macrina LC, Arrigo C. Passive range of motion characteristics in the overhead baseball pitcher and their implications for rehabilitation. Clin Orthop Relat Res. 2012;470(6):1586-1594.
23. Osbahr DC, Cannon DL, Speer KP. Retroversion of the humerus in the throwing shoulder of college baseball pitchers. Am J Sports Med. 2002;30(3):347-353.
24. Greenberg EM, Fernandez-Fernandez A, Lawrence JT, McClure P. The development of humeral retrotorsion and its relationship to throwing sports. Sports Health. 2015;7(6):489-496.
25. Jobe FW, Pink M. The athlete’s shoulder. J Hand Ther. 1994;7(2):107-110.
26. Montgomery WH 3rd, Jobe FW. Functional outcomes in athletes after modified anterior capsulolabral reconstruction. Am J Sports Med. 1994;22(3):352-358.
27. Jobe FW, Kvitne RS, Giangarra CE. Shoulder pain in the overhand or throwing athlete. The relationship of anterior instability and rotator cuff impingement. Orthop Rev. 1989;18(9):963-975.
28. Reinhold MM, Wilk KE, Dugas JR, Andrews JR. Chapter 11. Internal Impingement. In: Wilk K, Reinold MM, Andrews JR, eds. The Athlete’s Shoulder. 2nd ed. Philadelphia, PA: Churchill Livingstone Elsevier; 2009:126.
29. Kuhn JE, Lindholm SR, Huston LJ, Soslowsky LJ, Blasier RB. Failure of the biceps superior labral complex: a cadaveric biomechanical investigation comparing the late cocking and early deceleration positions of throwing. Arthroscopy. 2003;19(4):373-379.
30. Ciochon RL, Corruccini RS. The coraco-acromial ligament and projection index in man and other anthropoid primates. J Anat. 1977;124(Pt 3):627-632.
31. Moorman CT, Warren RF, Deng XH, Wickiewicz TL, Torzilli PA. Role of coracoacromial ligament and related structures in glenohumeral stability: a cadaveric study. J Surg Orthop Adv. 2012;21(4):210-217.
32. Su WR, Budoff JE, Luo ZP. The effect of coracoacromial ligament excision and acromioplasty on superior and anterosuperior glenohumeral stability. Arthroscopy. 2009;25(1):13-18.
33. Wellmann M, Petersen W, Zantop T, Schanz S, Raschke MJ, Hurschler C. Effect of coracoacromial ligament resection on glenohumeral stability under active muscle loading in an in vitro model. Arthroscopy. 2008;24(11):1258-1264.
34. Fagelman M, Sartori M, Freedman KB, Patwardhan AG, Carandang G, Marra G. Biomechanics of coracoacromial arch modification. J Shoulder Elbow Surg. 2007;16(1):101-116.
35. Bak K, Spring IB, Henderson IP. Re-formation of the coracoacromial ligament after open resection or arthroscopic release. J Shoulder Elbow Surg. 2000;9:289-293.
36. Levy O, Copeland SA. Regeneration of the coracoacromial ligament after acromioplasty and arthroscopic subacromial decompression. J Shoulder Elbow Surg. 2001;10(4):317-320.
37. Larson SG, Stern JT Jr. Role of supraspinatus in the quadrupedal locomotion of vervets (Cercopithecus aethiops): Implications for interpretation of humeral morphology. Am J Phys Anthropol. 1989;79(3):369-377.
38. Chansky HA, Iannotti JP. The vascularity of the rotator cuff. Clin Sports Med. 1991;10(4):807-822.
39. Klapper RC, Jobe FW, Matsuura P. Subscapularis muscle and its glenohumeral ligament-like bands. A histomorphologic study. Am J Sports Med. 1992;20(3):307-310.
40. Halder A, Zobitz ME, Schultz E, An KN. Structural properties of the subscapularis tendon. J Orthop Res. 2000;18(5):
829-834.
41. Elser F, Braun S, Dewing CB, Giphart JE, Millett PJ. Anatomy, function, injuries, and treatment of the long head of the biceps brachii tendon. Arthroscopy. 2011;27(4):581-592.
42. Pill SG, Walch G, Hawkins RJ, Kissenberth MJ. The role of the biceps tendon in massive rotator cuff tears. Instr Course Lect. 2012;61:113-120.
43. Krupp RJ, Kevern MA, Gaines MD, Kotara S, Singleton SB. Long head of the biceps tendon pain: differential diagnosis and treatment. J Orthop Sports Phys Ther. 2009;39(2):55-70.
44. Levy AS, Kelly BT, Lintner SA, Osbahr DC, Speer KP. Function of the long head of the biceps at the shoulder: electromyographic analysis. J Shoulder Elbow Surg. 2001;10(3):250-255.
45. Kuhn JE, Huston LJ, Soslowsky LJ, Shyr Y, Blasier RB. External rotation of the glenohumeral joint: ligament restraints and muscle effects in the neutral and abducted positions.
J Shoulder Elbow Surg. 2005;14(1 Suppl S):39S-48S.
46. McGarry MH, Nguyen ML, Quigley RJ, Hanypsiak B, Gupta R, Lee TQ. The effect of long and short head biceps loading on glenohumeral joint rotational range of motion and humeral head postion. Knee Surge Sports Traumatol Arthrosc. 2014 Sep 26. [Epub ahead of print].
47. Young RW. Evolution of the human hand: The role of throwing and clubbing. J Anat. 2003;202:165-174.
48. Calvin WH. Did throwing stones shape hominid brain evolution? Ethology and Sociobiology. 1982;3:115-124.
49. Fifer FC. The adoption of bipedalism by the hominids: A new hypothesis. Human Evolution. 1987;2(2):135-147.
50. Darlington PJ. Group selection, altruism, reinforcement, and throwing in human evolution. Proc Nat Acad Sci. 1973;72(9):3748-3752.
1. Darwin C. The Descent of Man, and Selection in Relation to Sex. 2nd ed. London, UK: John Murray; 1874:35.
2. Issac B. Throwing and human evolution. African Archeol Record. 1987;5:3-17.
3. Kirschann E. The human throw and a new model of hominid evolution (German). Homo. 1999;50(1):80-85.
4. Knusel CJ. The throwing hypothesis and hominid origins. Human Evolution. 1992;7(1):1-7.
5. Dunsworth H, Challis J, Walker A. The evolution of throwing: a new look at an old idea. Courier Forschungsinstitut Senckenberg. 2003;243:105-110.
6. Mayr E. Animal Species and Evolution. Cambridge, MA: Harvard University Press; 1963.
7. Tinbergen N. On the aims and methods of ethology. Zeitschrift für Tierpsychologie. 1963;20:410-433.
8. Fleisig GS, Andrews JR, Dillman CJ, Escamilla RF. Kinetics of baseball pitching with implications about injury mechanisms. Am J Sports Med. 1995;23(2):233-239.
9. Atwater AE. Biomechanics of overarm throwing movements and of throwing injuries. Exerc Sport Sci Rev. 1975;7:43-85.
10. Matsuo T, Escamila RF, Fleisig GS, Barrentine SW, Andrews JR. Comparison of kinematic and temporal parameters between different pitch velocity groups. J Appl Biomech. 2001;17:1-13.
11. Wang YT, Ford HT III, Ford HT Jr, Shin DM. Three-dimensional kinematic analysis of baseball pitching in acceleration phase. Percept Mot Skills. 1995;80:43-48.
12. Roach NT, Venkadesan M, Rainbow MJ, Lieberman DE. Elastic energy storage in the shoulder and the evolution of high-speed throwing in Homo. Nature. 2013;498(7455):483-486.
13. Fox JA, Cole BJ, Romeo AA, et al. Articular cartilage thickness of the humeral head: an anatomic study. Orthopedics. 2008;31(3):216.
14. Zumstein V, Kraljevic M, Conzen A, Hoechel S, Müller-Gerbl M. Thickness distribution of the glenohumeral joint cartilage: a quantitative study using computed tomography. Surg Radiol Anat. 2014;36(4):327-331.
15. Inman VT, Saunders M, Abbott LC. Observations on the function of the shoulder joint. J Bone Joint Surg Am. 1944;26:1-30.
16. O’Brien SJ, Schwartz RS, Warren RF, Torzilli PA. Capsular restraints to anterior posterior motion of the abducted shoulder: A biomechanical study. J Shoulder Elbow Surg. 1995;4(4):298-308.
17. Warner JJ, Deng XH, Warren RF, Torzilli PA. Static capsuloligamentous restraints to superior-inferior translation of the glenohumeral joint. Am J Sports Med. 1992;20(6):675-685.
18. Kuhn JE, Bey MJ, Huston LJ, Blasier RB, Soslowsky LJ. Ligamentous restraints to external rotation of the humerus in the late-cocking phase of throwing. A cadaveric biomechanical investigation. Am J Sports Med. 2000;28(2):200-205.
19. Bigliani LU, Codd TP, Connor PM, Levine WN, Littlefield MA, Hershon SJ. Shoulder motion and laxity in the professional baseball player. Am J Sports Med. 1997;25(5):609-613.
20. Borsa PA, Dover GC, Wilk KE, Reinold MM. Glenohumeral range of motion and stiffness in professional baseball pitchers. Med Sci Sports Exerc. 2006;38(1):21-26.
21. Hurd WJ, Kaplan KM, Eiattrache NS, Jobe FW, Morrey BF, Kaufman KR. A profile of glenohumeral internal and external rotation motion in the uninjured high school baseball pitcher, part I: motion. J Athl Train. 2011;46(3):282-288.
22. Wilk KE, Macrina LC, Arrigo C. Passive range of motion characteristics in the overhead baseball pitcher and their implications for rehabilitation. Clin Orthop Relat Res. 2012;470(6):1586-1594.
23. Osbahr DC, Cannon DL, Speer KP. Retroversion of the humerus in the throwing shoulder of college baseball pitchers. Am J Sports Med. 2002;30(3):347-353.
24. Greenberg EM, Fernandez-Fernandez A, Lawrence JT, McClure P. The development of humeral retrotorsion and its relationship to throwing sports. Sports Health. 2015;7(6):489-496.
25. Jobe FW, Pink M. The athlete’s shoulder. J Hand Ther. 1994;7(2):107-110.
26. Montgomery WH 3rd, Jobe FW. Functional outcomes in athletes after modified anterior capsulolabral reconstruction. Am J Sports Med. 1994;22(3):352-358.
27. Jobe FW, Kvitne RS, Giangarra CE. Shoulder pain in the overhand or throwing athlete. The relationship of anterior instability and rotator cuff impingement. Orthop Rev. 1989;18(9):963-975.
28. Reinhold MM, Wilk KE, Dugas JR, Andrews JR. Chapter 11. Internal Impingement. In: Wilk K, Reinold MM, Andrews JR, eds. The Athlete’s Shoulder. 2nd ed. Philadelphia, PA: Churchill Livingstone Elsevier; 2009:126.
29. Kuhn JE, Lindholm SR, Huston LJ, Soslowsky LJ, Blasier RB. Failure of the biceps superior labral complex: a cadaveric biomechanical investigation comparing the late cocking and early deceleration positions of throwing. Arthroscopy. 2003;19(4):373-379.
30. Ciochon RL, Corruccini RS. The coraco-acromial ligament and projection index in man and other anthropoid primates. J Anat. 1977;124(Pt 3):627-632.
31. Moorman CT, Warren RF, Deng XH, Wickiewicz TL, Torzilli PA. Role of coracoacromial ligament and related structures in glenohumeral stability: a cadaveric study. J Surg Orthop Adv. 2012;21(4):210-217.
32. Su WR, Budoff JE, Luo ZP. The effect of coracoacromial ligament excision and acromioplasty on superior and anterosuperior glenohumeral stability. Arthroscopy. 2009;25(1):13-18.
33. Wellmann M, Petersen W, Zantop T, Schanz S, Raschke MJ, Hurschler C. Effect of coracoacromial ligament resection on glenohumeral stability under active muscle loading in an in vitro model. Arthroscopy. 2008;24(11):1258-1264.
34. Fagelman M, Sartori M, Freedman KB, Patwardhan AG, Carandang G, Marra G. Biomechanics of coracoacromial arch modification. J Shoulder Elbow Surg. 2007;16(1):101-116.
35. Bak K, Spring IB, Henderson IP. Re-formation of the coracoacromial ligament after open resection or arthroscopic release. J Shoulder Elbow Surg. 2000;9:289-293.
36. Levy O, Copeland SA. Regeneration of the coracoacromial ligament after acromioplasty and arthroscopic subacromial decompression. J Shoulder Elbow Surg. 2001;10(4):317-320.
37. Larson SG, Stern JT Jr. Role of supraspinatus in the quadrupedal locomotion of vervets (Cercopithecus aethiops): Implications for interpretation of humeral morphology. Am J Phys Anthropol. 1989;79(3):369-377.
38. Chansky HA, Iannotti JP. The vascularity of the rotator cuff. Clin Sports Med. 1991;10(4):807-822.
39. Klapper RC, Jobe FW, Matsuura P. Subscapularis muscle and its glenohumeral ligament-like bands. A histomorphologic study. Am J Sports Med. 1992;20(3):307-310.
40. Halder A, Zobitz ME, Schultz E, An KN. Structural properties of the subscapularis tendon. J Orthop Res. 2000;18(5):
829-834.
41. Elser F, Braun S, Dewing CB, Giphart JE, Millett PJ. Anatomy, function, injuries, and treatment of the long head of the biceps brachii tendon. Arthroscopy. 2011;27(4):581-592.
42. Pill SG, Walch G, Hawkins RJ, Kissenberth MJ. The role of the biceps tendon in massive rotator cuff tears. Instr Course Lect. 2012;61:113-120.
43. Krupp RJ, Kevern MA, Gaines MD, Kotara S, Singleton SB. Long head of the biceps tendon pain: differential diagnosis and treatment. J Orthop Sports Phys Ther. 2009;39(2):55-70.
44. Levy AS, Kelly BT, Lintner SA, Osbahr DC, Speer KP. Function of the long head of the biceps at the shoulder: electromyographic analysis. J Shoulder Elbow Surg. 2001;10(3):250-255.
45. Kuhn JE, Huston LJ, Soslowsky LJ, Shyr Y, Blasier RB. External rotation of the glenohumeral joint: ligament restraints and muscle effects in the neutral and abducted positions.
J Shoulder Elbow Surg. 2005;14(1 Suppl S):39S-48S.
46. McGarry MH, Nguyen ML, Quigley RJ, Hanypsiak B, Gupta R, Lee TQ. The effect of long and short head biceps loading on glenohumeral joint rotational range of motion and humeral head postion. Knee Surge Sports Traumatol Arthrosc. 2014 Sep 26. [Epub ahead of print].
47. Young RW. Evolution of the human hand: The role of throwing and clubbing. J Anat. 2003;202:165-174.
48. Calvin WH. Did throwing stones shape hominid brain evolution? Ethology and Sociobiology. 1982;3:115-124.
49. Fifer FC. The adoption of bipedalism by the hominids: A new hypothesis. Human Evolution. 1987;2(2):135-147.
50. Darlington PJ. Group selection, altruism, reinforcement, and throwing in human evolution. Proc Nat Acad Sci. 1973;72(9):3748-3752.